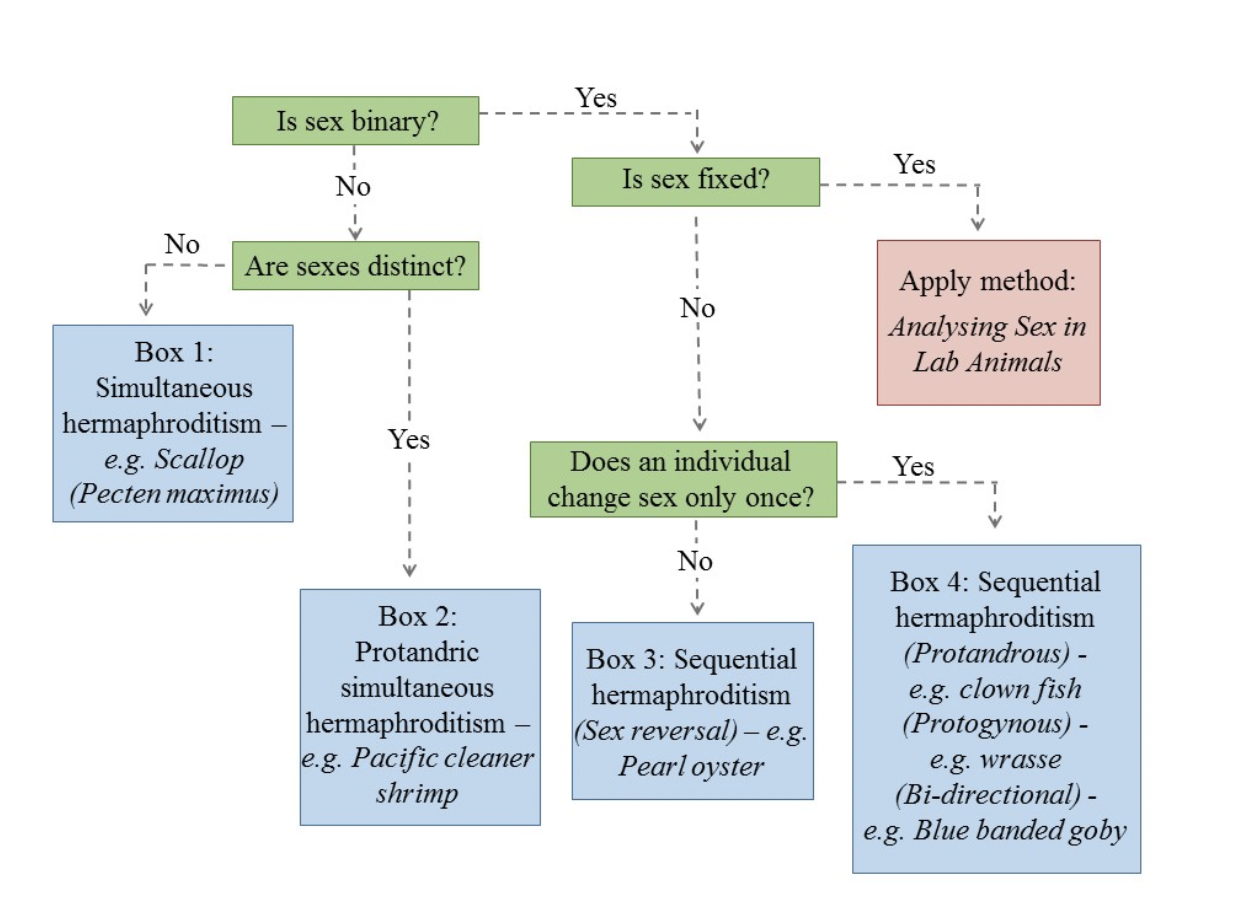
Box 1– Simultaneous hermaphrodites contain both male and female gametes in the same adult at the same time. A familiar example is the king scallop, Pecten maximus, where the bi-colored roe results from oocytes (orange tissue) and spermatozoa (cream tissue) (Paz et al., 2005). In simultaneous hermaphrodites, the proportion of reproductive investment allocated to gametes of each sex can change during an individual’s life (Janicke et al., 2013; Leonard, 2013); thus the impact of sex on whole organism energetics will depend on the balance of male and female tissue within an individual. Sex analysis with simultaneous hermaphrodites must consider the proportion of tissues with defined sex within an individual, in addition to how tissue sex determines the response of the whole organism.
 King scallop (Pecten maximus). Bi-colored roe indicate female (orange) and male (ream) gametes. Image rights CC0 1.0 Universal.
King scallop (Pecten maximus). Bi-colored roe indicate female (orange) and male (ream) gametes. Image rights CC0 1.0 Universal.
Box 2– Pacific cleaner shrimp are protandric simultaneous hermaphrodites—they develop as males and later change sex to simultaneous hermaphrodites (Fiedler, et al., 2010). Following sex change, individuals have both male and female tissues in their gonads and produce both gametes. Sex analysis in such organisms must include the impact of sex change processes on the organism’s response to environmental stress. To understand how the response of males and hermaphrodites differs when exposed to stress, researchers must account for the impact of male and female tissues, gametes and cellular processes within hermaphrodites, and how these interact within the whole organism. In addition, as with all simultaneous hermaphrodites, the comparison should account for the energy reserves and tissue volume an individual allocates to the production of each sex tissue proportionally (Janicke et al., 2013; Leonard, 2013), where possible (i.e. how female are simultaneous hermaphrodites?).
Box 3– Pearl oysters develop primarily as male and later change to female (Adzigbli, et al., 2019). However, sex reversal can subsequently occur as a result of environmental conditions, with the sex ratio linked to season (Southgate & Lucas, 2011). Moreover, in this species, the production and quality of pearls (Adzigbli, et al., 2019), and thus the economic value of the species, depends on sex. Males produce higher quality pearls. Sex analysis in species like the pearl oyster, in which multiple sex reversals are possible, must investigate sex-determining mechanisms and how changing sex, possibly multiple times, impacts an individual’s performance, such as pearl production, growth or sexual reproduction.
Box 4 – Sequential hermaphroditism can be protandrous (male to female), as in the case of clownfish (Casas et al., 2016); protogynous (female to male), as exhibited in wrasse species (Munday et al., 2006); or bi-directional (sex reversal in one direction irrespective of initial sex), as demonstrated in bluebanded gobies (Rodgers et al., 2007). While sexes are distinct and, following sex change, individuals become fully functional as the opposite sex, the sex change process within an individual occurs over multiple time frames (Lamm et al., 2015). In bluehead wrasse, the removal of the dominant terminal male from a reef induces large females to change sex (Lamm et al., 2015). However, while the behavior of this female changes to that typical of a male within minutes, gonadal sex change occurs over 1 to 2 weeks (Lamm et al., 2015). An area yet to be explored with respect to sex analysis and climate change is how an individual will respond to stress during this sex change process.
A) Protandric hermaphrodite Red Sea anemone fish
B) Protogynous hermaphrodite bluehead wrasse
C) Bi-directional hermaphrodite bluebanded goby Pacific cleaner shrimp (Lysmata amboinensis) are protrandric simultaneous hermaphrodites. They mature as male and later change sex to hermaphrodite. Photo credit: Lonnie Huffman.
Pacific cleaner shrimp (Lysmata amboinensis) are protrandric simultaneous hermaphrodites. They mature as male and later change sex to hermaphrodite. Photo credit: Lonnie Huffman.
https://creativecommons.org/licenses/by/3.0/deed.en.

 Sequentially hermaphroditic fish.
Sequentially hermaphroditic fish.
(Amphiprion bicinctus), photo credit: Mal B
https://creativecommons.org/licenses/by-nd/2.0/
(Thalassoma bifasciatum)photo credit: Kevin Bryant
https://creativecommons.org/licenses/by-nd/2.0/
(Lythrypnus dally), photo credit: Richard Ling
https://creativecommons.org/licenses/by-nc-nd/2.0/
Works Cited
Adzigbli, L., Wang, Z., Lai, Z., Li, J., &Deng, Y. (2019). Sex determination in pearl oyster: a mini review. Aquaculture Reports, 15. https://doi.org/10.1016/j.aqrep.2019.100214
Casas, L., Saborido-Rey, F., Ryu, T., Michell, C., Ravasi, T., & Irigoien, X. (2016). Sex change in clownfish: molecular insights from transcriptome analysis. Scientific reports, 6. https://doi.org/10.1038/srep35461 Fiedler, G. C., Rhyne, A. L., Segawa, R., Aotsuka, T., & Schizas, N. V. (2010). The evolution of euhermaphroditism in caridean shrimps: a molecular perspective of sexual systems and systematics. BMC evolutionary biology, 10(1), 297. Janicke, T., Marie‐Orleach, L., De Mulder, K., Berezikov, E., Ladurner, P., Vizoso, D. B., & Schärer, L. (2013). Sex allocation adjustment to mating group size in a simultaneous hermaphrodite. Evolution, 67(11), 3233-3242. Lamm, M. S., Liu, H., Gemmell, N. J., & Godwin, J. R. (2015). The need for speed: neuroendocrine regulation of socially-controlled sex change. Integrative and Comparative Biology, 55(2), 307-322. Leonard, J. L. (2013). Williams' paradox and the role of phenotypic plasticity in sexual systems. Integrative and comparative biology, 53(4), 671-688. Munday, P. L., Wilson White, J., & Warner, R. R. (2006). A social basis for the development of primary males in a sex-changing fish. Proceedings of the Royal Society B: Biological Sciences, 273(1603), 2845-2851. Paz, M., Torrado, M., Korochkin, L. I. and Mikhailov, A. T. (2005). Esterase-like and fibronectin-like polypeptides share similar sex-cell-biased patterns in the gonad of hermaphroditic and gonochoric species of bivalve mollusks. Cell and tissue research, 322(3), 475-489. Rodgers, E. W., Earley, R. L., & Grober, M. S. (2007). Social status determines sexual phenotype in the bi‐directional sex changing bluebanded goby Lythrypnus dalli. Journal of Fish Biology, 70(6), 1660-1668. Southgate, P., & Lucas, J. (Eds.). (2011). The Pearl Oyster. Oxford: Elsevier.